


| name | Amanita sp-AZ16 | ||||||||
| author | Tulloss | ||||||||
| name status | cryptonomen temporarium | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| intro |
Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is based on original research of R. E. Tulloss. | ||||||||
| pileus | 52 - 66 mm wide, yellow (3A3-4) with disc dingy in one specimen, more pallid at margin (cream yellow to pale yellow (between 5Y 8/4 and 5Y 8/6)), somewhat irregularly hemispheric with flattened disc, with margin slightly incurved, tacky, matt to subshiny; context pale yellow, becoming somewhat paler toward lamellae, unchanging when cut or bruised, 5.5 - 6.5 mm thick, thinning evenly to margin; margin striate (0.5 - 1.5R), nonappendiculate; universal veil as small patches and warts about 1* mm thick, white, soft, with some fibrils on surface, submembranous to subfelted, detersile, darkening somewhat with exposure (off-white, then grayish or pale brownish gray). | ||||||||
| lamellae | free, without decurrent line on stipe apex, crowded, pale cream in mass, pale cream to off-white in side view, 2.5 - 4.5 mm broad; lamellulae truncate, unevenly distributed, of diverse lengths, plentiful. | ||||||||
| stipe | 39 - 73 × 9.5 - 11 mm, white, browning from handling, narrowing upward, not flaring at apex, finely pruinose above, faintly silky-fibrillose below, stipe splitting into recurved scales; bulb 18 - 23 × 18 - 24 mm, subnapiform, sometimes slightly flattened; context pale yellow above to whitish below, not rapidly changing when cut or bruised, orangy tan in old and extensive damaged areas, firmly stuffed with white material, with central cylinder 4 mm wide; exannulate in immature specimens; universal veil as thin, white collar—collapsed limb to subocreate. | ||||||||
| odor/taste | Odor indistinct. Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
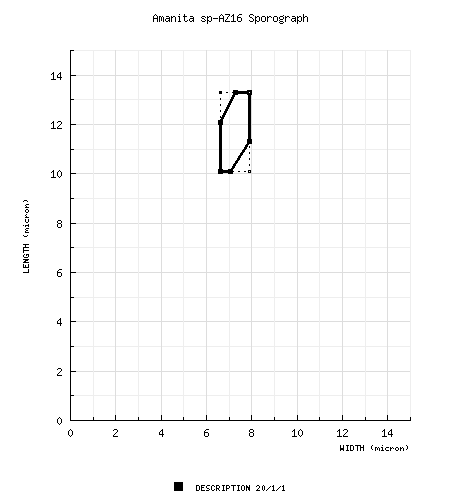
| basidiospores | BASIDIOSPORES: [20/1/1] (9.9-) 10.1 - 13.3 (-14.3) × (6.0-) 6.6 - 7.9 (-8.2) µm, (L = 11.9 µm; W = 7.3 µm; Q = 1.43 - 1.83 (-1.96); Q 1.62), ??, ??, smooth, thin-walled, inamyloid, ellipsoid to elongate, adaxially flattened, sometimes swollen at one end;apiculus sublateral, cylindric, prominent; contents ??; color in deposit not recorded. | ||||||||
| ecology | Gregarious at 1535 m elev. In alluvial soil (sandy loam) under Quercus reticulata in type B habitat. | ||||||||
| material examined | U.S.A.: ARIZONA—Cochise Co. - CMP site #27, 18.viii.1992 R. L. Homola s.n. [Tulloss 8-18-92-A; CMP0570] (RET ??). | ||||||||
| discussion |
The most similar spores in the currently described North American taxa suspected from the region of study belong to A. velatipes (Q' = 1.44), but this entity has a thick, cothurnate rim on the top of the bulb and a persistent partial veil. Amanita xylinivolva has spores with Q’ = 1.10, a partial veil that is present in immature material, a broadly conic shape in the immature pileus, a context that is basically white in both pileus and stipe, and a volva represented by rather large, membranous to submembranous limbs. The bulbs of the specimens of the sole known collection were badly decayed. It was not possible to further expand the pileus of the largest and most mature. The stipes had surface pitting and recurved scales because of insect damage. These same insects may have eaten the partial veils. After measuring the spores, the possibility of some “early sporulation” abnormality should be carefully checked. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.
